






INTRODUCTION
The craniovertebral junction (CVJ) is a highly complex functional unit consisting of the base of the occipital bone, the atlas (C1), the axis (C2) and the soft tissues around these skeletal structures [1,2]. It provides complex motion of upper cervical region and protects the crucial aera of the brainstem and upper cervical spinal cord. CVJ skeletal malformation can result in structural instability causing spectrum of neurological deficits including sensory and motion dysfunction, or even life-threatening dyspnea and cardiac arrest [3]. The spectrum of CVJ malformation includes atlantooccipital fusion, atlantoaxial dislocation, basilar invagination, platybasia, and vertebral artery variations [4]. It is intuitive to attribute the etiology of CVJ malformation to disturbance of embryonic development. Both genetic and environmental factors can be involved. Homeobox (HOX) genes are pivotal genetic regulators of embryonic axial development [2]. For instance, inactivating Hoxd3 in mice causes abnormal fusion of the atlas vertebra and the occiput, which is homologous to atlantooccipital fusion in human [5]. However, the genetic causes for the CVJ malformation were largely unclear.
Several studies suggested genetic predisposition underlying CVJ malformation. CVJ malformation has been reported to be much more common in Down syndrome (MIM: 190685) [6-8], Mucopolysaccharidosis type IVA (MIM: 253000) [9], and Wiedemann-Steiner syndrome (MIM: 605130) [10]. Sadler et al. [11] found that chromodomain (CHD) genes were associated with Chiari I malformation. Chauhan et al. [12] reported mutations in the FBN1 gene may play a part in CVJ malformation. In the previous study, we performed exome sequencing on 92 sporadic CVJ malformation patients and found damaging mutations of the BEST1 gene were significantly enriched in the patient group (p = 5.8× 10-6) [13]. Variants in BEST1 are associated with autosomal dominant vitelliform macular dystrophy-2 (VMD2 [MIM: 153700]) [14,15]. Inhibition of BEST1 resulted in an enhanced neuronal excitability in the cerebellum [16]. These studies indicated a disease-causing role of BEST1 gene mutations for eyeball, central nervous system, and maybe skeletal system disorders.
Given that the BEST1 gene is highly conserved in zebrafish and human, and zebrafish has been proven to be suitable for the study of human skeletal disease [17-19], we designed best1 morpholinos (MOs) and performed knockdown studies to investigate the role of best1 gene in early zebrafish development.
MATERIALS AND METHODS
1. Zebrafish Care and Maintenance
Adult wild-type AB strain zebrafish were maintained at 28.5°C on a 14-hour light/10-hour dark cycle [20]. Five to 6 pairs of zebrafish were set up for natural mating every time. On average, 200–300 embryos were generated. Embryos were maintained at 28.5°C in fish water (0.2% Instant Ocean Salt in deionized water). The embryos were washed and staged according to Kimmel et al. [21]. The establishment and characterization of HuC-EGFP transgenic lines has been described elsewhere [22]. The zebrafish facility at SMOC (Shanghai Model Organisms Center, Inc., Shanghai, China) is accredited by the AAALAC (Association for Assessment and Accreditation of Laboratory Animal Care) International.
2. Zebrafish Microinjections and Phenotyping
It is imperative to identify a sequence within the best1 gene that is pivotal for its physiological function. We also avoid regions with high sequence homology to other genes, to reduce the chance of off-target effects. Gene Tools, LLC (http://www.gene-tools.com/) designed the MOs. Antisense MOs (GeneTools) were microinjected into fertilized one-cell stage embryos according to standard protocols [23]. The sequences of the best1 translation-blocking and splice-blocking MOs were 5′-CTGCGGGAGAGTCCAAAACACTTCA-3′ (ATG-MO) and 5′-GAGTACGTCACCGTCATCACTG-3′ (I3E4-MO), respectively. The sequence for the standard control-MO was 5′-CCTCTTACCTCAGTTACAATTTATA-3′ (Gene Tools). The amount of the MOs used for injection was as follows: Control-MO and I3E4-MO, 4 ng per embryo; ATG-MO, 4 ng per embryo. Primers spanning best1 exon 3 (forward primer: 5′-GCAGATGAAGGAGCCAGACT-3′) and exon 6 (reverse primer: 5′-AAACTGCCGACCAATCAGAC-3′) were used for reverse transcription-polymerase chain reaction (RT-PCR) analysis for confirmation of the efficacy of the I3E4-MO. The primer ef1α sequences used as the internal control were 5′-GGAAATTCGAGACCAGCAAATAC-3′ (forward) and 5′-GATACCAGCCTCAAACTCACC-3′ (reverse). At 2-dpf and 5-dpf, embryos were anesthetized with 0.016% MS-222 (tricaine methanesulfonate, Sigma-Aldrich, St. Louis, MO, USA). Zebrafish were then oriented on lateral side (anterior, left; posterior, right; dorsal, top), and mounted with 3% methylcellulose in a depression slide for observation by fluorescence microscopy. The phenotypes of gross morphology were quantified analyzed.
3. Quantitative Real-Time PCR
Total RNA was extracted from 20 to 30 embryos per group with Trizol (Roche, Basel, Switzerland) according to the manufacturer’s instructions. RNA was reverse transcribed using the the PrimeScript RT reagent Kit with gDNA Eraser (Takara, Tokyo, Japan). Quantification of gene expression was performed in triplicates using Bio-rad iQ SYBR Green Supermix (Bio-Rad Laboratories Inc., Hercules, CO, USA) with detection on the Realplex system (Eppendorf Co., Enfield, CT, USA). Relative gene expression quantification was based on the comparative threshold cycle method (2−ΔΔCt) using ef1α as endogenous control gene. Primer sequences are listed in Supplementary Table 1.
4. Calcein Staining
To evaluate craniofacial structure formation in zebrafish, fertilized one-cell wild-type AB embryos were injected with best1-MO and control-MO. At 6-dpf, zebrafish were washed with fish water 3 times and immersed in 0.2% Calcein solution for 10 minutes. Then, zebrafish were rinsed thoroughly in fish water 3 times (5 minutes for each time) and anaesthetized with 0.016% MS-222 (tricaine methanesulfonate, Sigma-Aldrich). Zebrafish were then oriented on lateral side and ventral side, and mounted with 3% methylcellulose (Sigma-Aldrich) in a depression slide for observation by fluorescence microscopy.
5. Image Acquisition
Embryos and larvae were analyzed with Nikon SMZ 18 Fluorescence microscope and subsequently photographed with digital cameras. A subset of images was adjusted for levels, brightness, contrast, hue and saturation with Adobe Photoshop 7.0 software (Adobe, San Jose, CA, USA) to optimally visualize the expression patterns. Quantitative image analyses were processed using image based morphometric analysis (NIS-Elements D4.6, Nikon, Tokyo, Japan) and ImageJ software (U.S. National Institutes of Health, Bethesda, MD, USA; http://rsbweb.nih.gov/ij/). 10 animals were including in group and average signal per animal was calculated.
6. Statistical Analysis
All data are presented as mean ± standard error of the mean (SEM). Statistical analysis and graphical representation of the data were performed using GraphPad Prism 5.0 (GraphPad Software, San Diego, CA, USA). Statistical significance was performed using a Student t-test, analysis of variance or χ2 test as appropriate.
RESULTS
1. Effectiveness of best1 I3E4-MO
Customized MOs were microinjected into one-cell stage embryos (Supplementary Fig. 1A). Two specific antisense MOs targeting zebrafish best1 gene were designed to prevent either the translation of best1 mRNA (ATG-MO) or proper splicing of intron3 and exon4 (I3E4-MO) of best1 pre-mRNA (Supplementary Fig. 1B). We confirmed that I3E4-MO prevented proper splicing of intron3 at 30-hpf by RT-PCR. The additional band indicated that the splicing between intron 3 and exon 4 was altered with inserting part of intron3 (Supplementary Fig. 1C, lane 2). In addition, the expression of house-keeping genes ef1α was not influenced by both control-MO and I3E4-MO (Supplementary Fig. 1C, lanes 5 and 6). The expression level of wildtype best1 mRNA in I3E4-MO injected embryos was significantly lower than that of the control (p < 0.0001; Supplementary Fig. 1D) with quantitative RT-PCR. This demonstrated the efficacy of I3E4-MO to knocked down best1 gene.
2. Knockdown of best1 Causes Potent Developmental Defects in Zebrafish
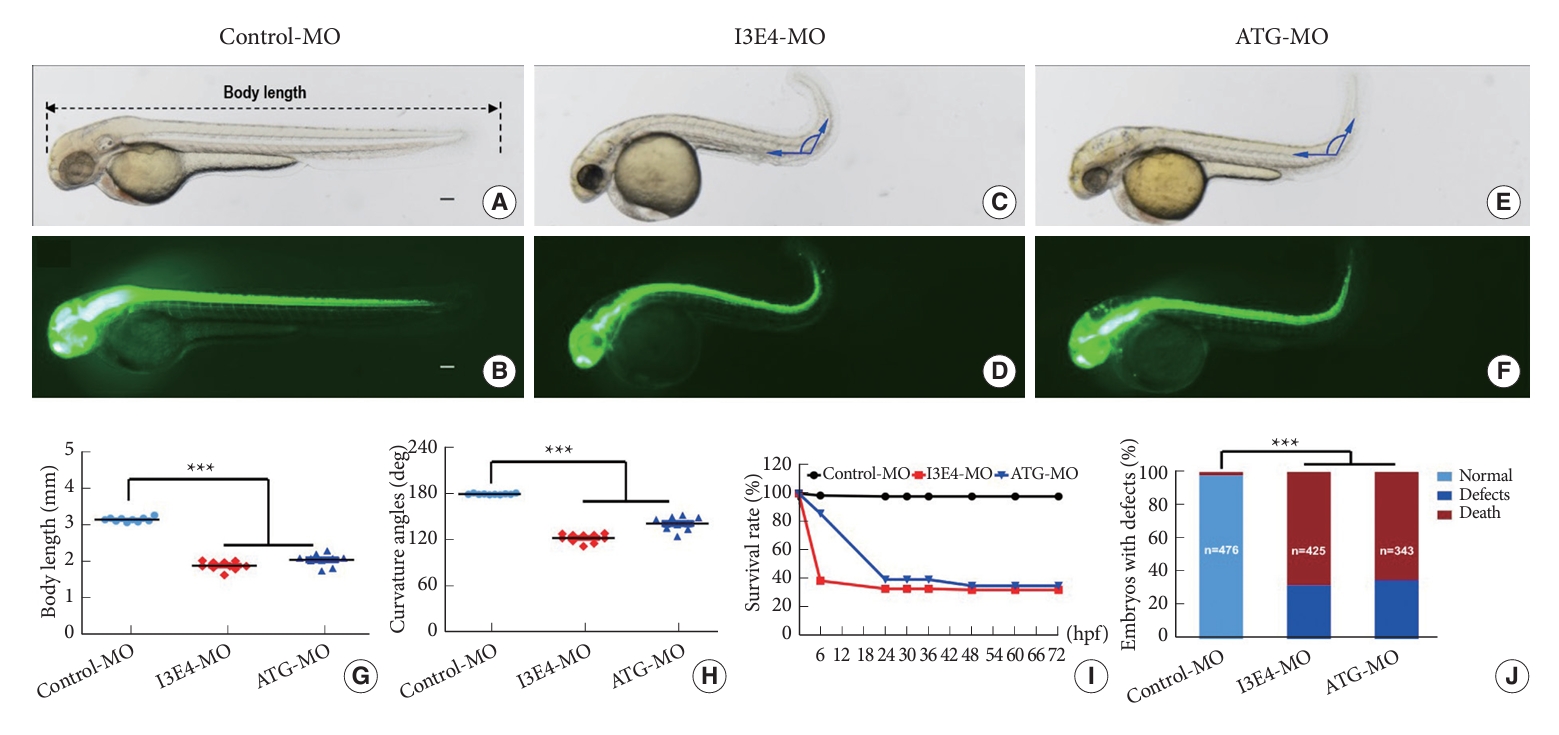
We used transgenic (Tg) zebrafish (HuC: EGFP transgenic line) to observe the phenotypes of zebrafish embryos after best1 knockdown by injection of ATG-MO and I3E4-MO. To increase the objectivity of the measurements, we performed the quantification process in a blinded manner. This approach concealed the experimental groups during the measurement procedure, avoiding potential bias in the evaluation of phenotypic changes. Compared to control-MO injected embryos (Fig. 1A and B), both I3E4-MO injected (Fig. 1C and D) and ATG-MO injected (Fig. 1E and F) embryos exhibited significantly reduced body length (p < 0.0001) (Fig. 1G) and apparent curved body axis (p < 0.0001) (Fig. 1D, F, and H; Supplementary Fig. 2). The survival rate of best1 knockdown embryos was decreased (Fig. 1I) and the percentage of live embryos with aforementioned gross defects was significantly higher in the best1 knockdown group versus the control group (p < 0.0001) (Fig. 1J).
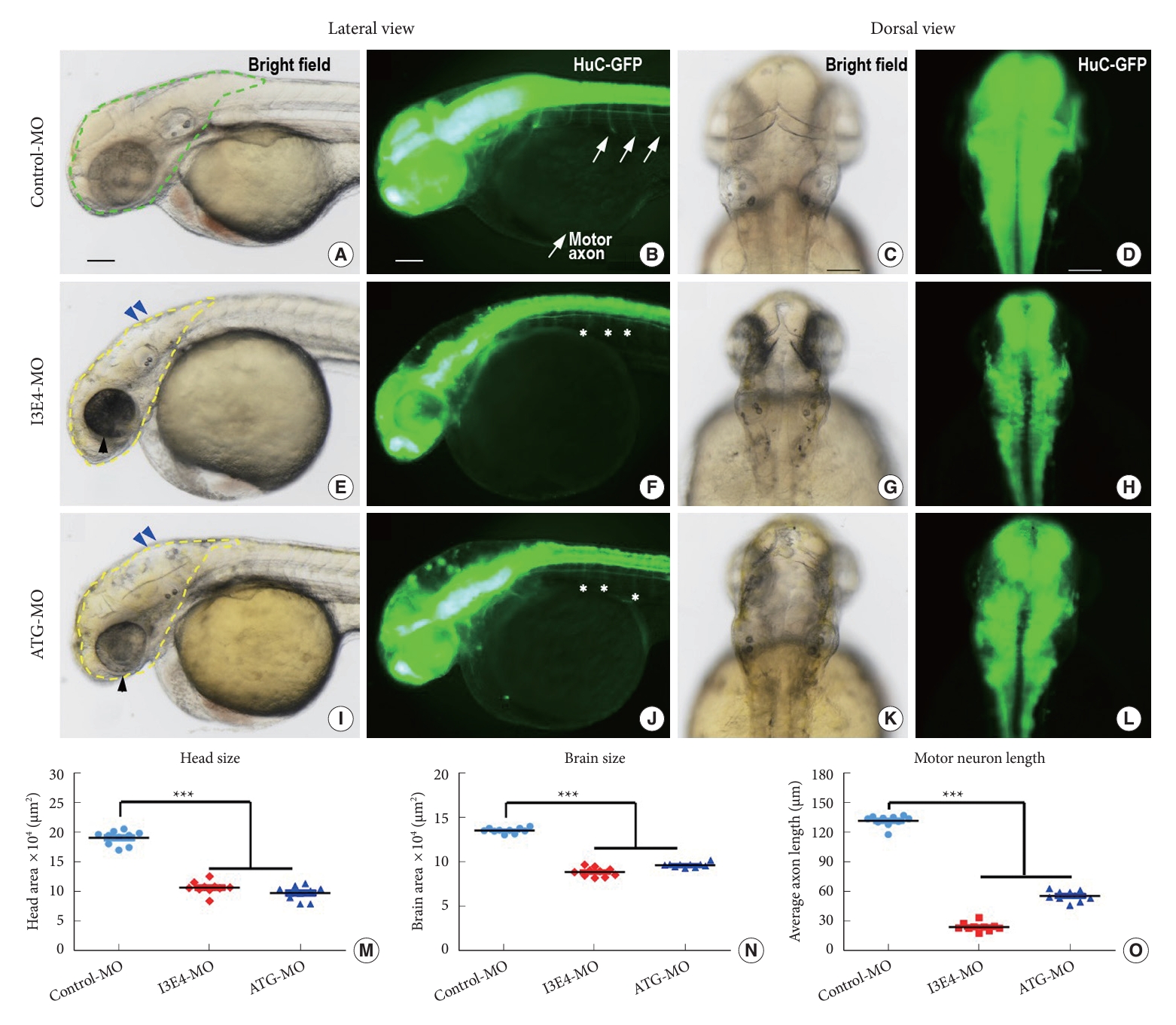
Knockdown of best1 caused several developmental abnormalities in the nervous system, including defects in head and brain patterning (Fig. 2A, E, and I, denoted by dotted lines and blue arrowheads) and defective neuronal outgrowth (Fig. 2B, F, and J, arrows in control embryo indicate normal motor axons; asterisks in I3E4-MO and ATG-MO embryos showed no outgrowth of axons). Quantitative morphometric analysis revealed substantially decreased head and brain sizes (p < 0.0001) (Fig. 2M and N) and markedly shortened motor neuron length (p < 0.0001) (Fig. 2O) in I3E4-MO embryos compared to controls. Knockdown of best1 also resulted in reduced eye size of zebrafish (Fig. 2E and I, marked by black arrowheads).
Taken together, these results demonstrate that the knockdown of best1 in zebrafish embryos can lead to shorter body length, curved body axis, low survival rate, microcephaly, small eyes, reduced head and brain size, and impaired neuronal outgrowth.
3. Knockdown of best1 Causes Craniofacial and Vertebral Skeletal Ossification Defect
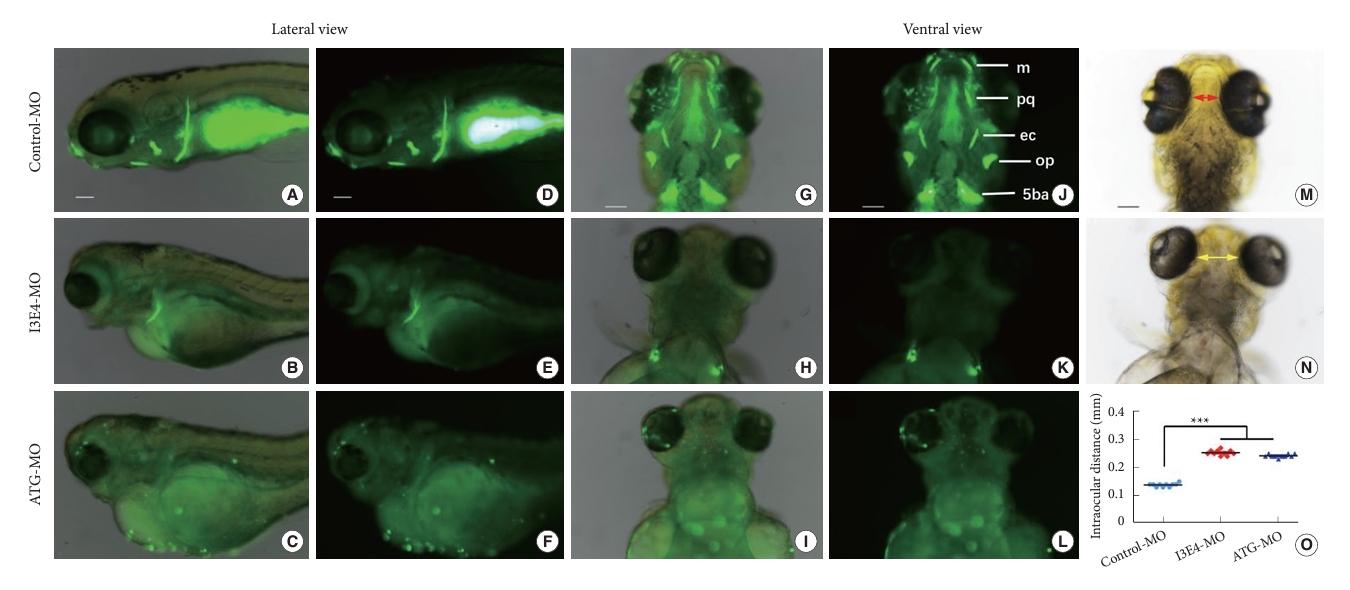
By application of the fluorescent dye Calcein to label the calcified matrix of the skeleton, a significant reduction of craniofacial skeletal ossification was found in best1 knockdown zebrafish (6 days post fertilization, 6 dpf) compared to that of control (Fig. 3). In the control embryos, 5 pairs of craniofacial bone structures were nicely labeled in both lateral (Fig. 3A and D) and ventral (Fig. 3G and J). However, only the fifth branchial arch (5ba) was faintly labeled in the I3E4-MO embryos (Fig. 3B, E, H, and K), and no calcified bone structures were identified in the ATG-MO embryos (Fig. 3C, F, I, and L). Impaired craniofacial development also led to increased interorbital distance in best1 knockdown embryos (p < 0.0001) (Fig. 3M–O).
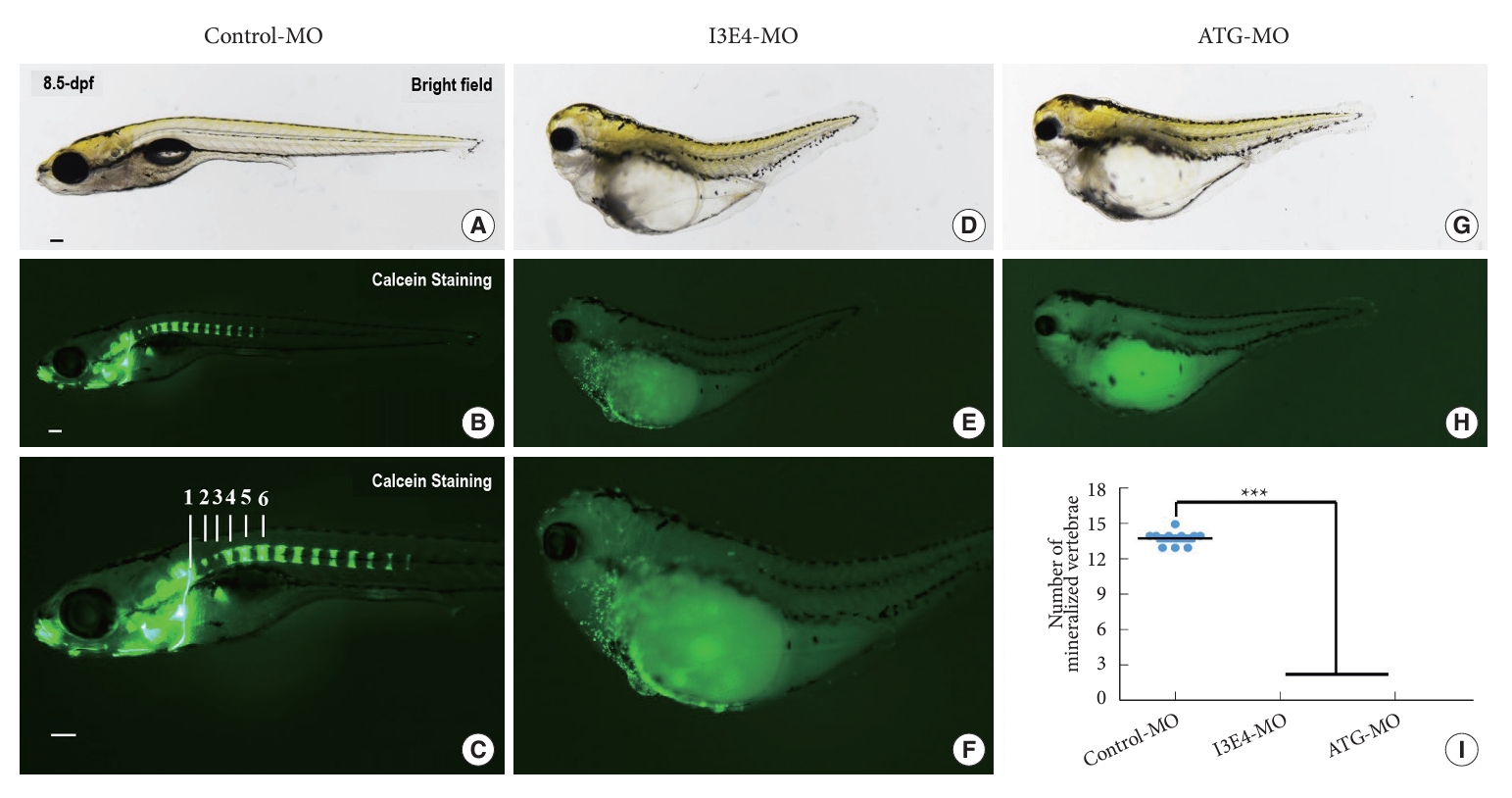
Furthermore, we administrated Calcein at 8.5 dpf to visualize calcified vertebrae of zebrafish embryos. On average, 14 vertebral segments could be clearly identified in the control group (Fig. 4A–C). Whereas zero vertebrae could be distinguished in best1 knockdown embryos (Fig. 4D–H). There is a significant difference among the groups (p < 0.0001) (Fig. 4I).
These findings indicated that best1 might play a critical role in the early skeletal development of zebrafish.
DISCUSSION
Congenital CVJ skeletal malformation is a rare disease that can lead to disastrous neurological deficits in patients. The etiology of congenital CVJ malformation is still undefined. In our previous study, 5 loss-of-function or damaging missense were detected out of 92 congenital CVJ skeletal malformation patients (carrier rate = 5.43%), which is significantly enriched in the patient group compared with in-house control (p = 5.77×10–6; odds ratio, 27.74; 95% confidence interval, 7.15–94.25) [13]. In this study, we designed MOs targeting zebrafish best1 to knockdown best1 expression and investigated the corresponding phenotypes.
First, the knockdown of best1 in zebrafish embryos caused smaller eyes than control. Microphthalmia (axial length≤20 mm) has been reported in multiple VMD2 patients [24,25]. It has been reported that BEST1 played an essential role in ocular patterning, maintenance of retinal pigment epithelium (RPE) cells, and normal growth of the eye [26]. Bestrophin is a 585 amino acid transmembrane protein acting as an oligomeric chloride channel that regulates calcium channels for calcium and transmembrane potential homeostasis [27]. Bestrophin mutations alter chloride ion-related conductance across the RPE cell membrane, and abnormal channel function may account for the abnormal electro-oculogram seen in patients with Best disease. Our results in zebrafish agreed with that in VMD2 patients.
Second, the knockdown of best1 in zebrafish embryos resulted in multiple skeletal defects, including shorter body length, curved body axis, microcephaly, and impaired ossification of craniofacial and vertebral bone. Early zebrafish skeleton patterning is regulated by complex signaling pathways, including hox, fgf, shh, tbx, hmx, runx2, bmp, and others [28,29]. Intriguingly, skeleton malformations can be associated with eye and ear disorders. In human Oculoauricular Syndrome (MIM 612109), patients manifest with complex ocular anomalies, including microphthalmia, dysplastic ears with abnormal external ear cartilage, as well as spina bifida occulta and platybasia [30]. In mice, mutations in Mitf gene can cause microphthalmia, deafness and osteopetrosis [31]. BEST1 is regulated by MITF [32], these results imply that BEST1 gene plays an important role in eye development and skeletal ossification. In addition, BEST1 interacts with ANO6, another calcium-dependent transmembrane channel, which is crucial for apatite deposition in bone mineralization [33]. ANO6 activates NCX1, a calcium channel in osteoblasts, to transport calcium to the extracellular matrix for bone formation. Ano6-/- mouse embryos (E15.5) also show delayed bone mineralization [34], which is highly consistent with our zebrafish results.
Third, best1 knockdown caused microcephaly, reduced brain size, and impaired neuronal outgrowth in zebrafish embryos. Woo et al. [16] prepared Best1 knockout mice to investigate tonic inhibition in the brain and found that inhibition of Best1 resulted in enhanced neuronal excitability in the cerebellum. Apparently Bestrophins distribute and function as ion channels in the central nervous system [27]. However, no studies have yet reported these neurological developmental defects.
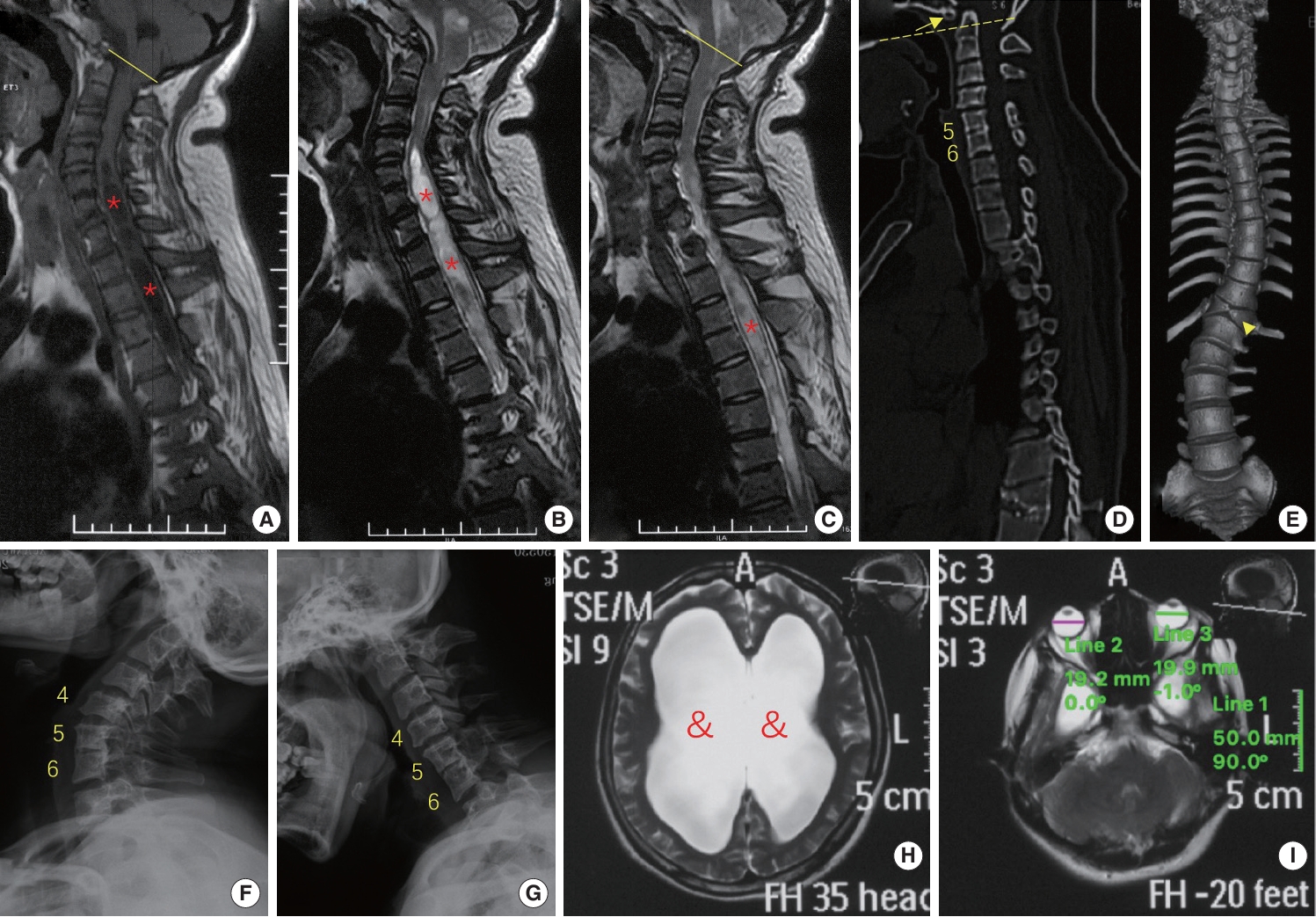
Intriguingly, the patient in our cohort with a premature stop codon of BEST1 gene (p.S79Ffs*153) did present with ophthalmic, spinal and neurological developmental defects (Fig. 5). This is a 27-year-old female patient suffering with decreased sensation on all limbs and weakness of bilateral lower limbs for one year. Physical examination revealed increased tendon reflexes and positive Hoffmann and Babinski signs. Imaging workup found Chiari malformation with syringomyelia and hydrocephalus, multiple spinal anomalies (including assimilation of atlas, basilar invagination, fusion of C5 and C6 vertebrae, butterfly vertebra at T12 and scoliosis) and microphthalmia. She denied decreased visual acuity, which is a later-onset symptom of VMD2, and refused a fundus examination. The specific genetic mutation identified in the patient was a premature stop codon, which could result in truncation of the Bestrophin-1 protein, affecting its normal structure and function. Interestingly, this mutation’s impact in humans is similar to that of MO, which knocks down the expression of the best1 gene in zebrafish. Notably, this premature stop codon mutation was identified in a CVJ malformation patient who also presented comorbidities such as microphthalmia, neurological defects, and multiple skeletal malformations, particularly along the spine. Given the homology between zebrafish and humans, it is hypothesized that decreased BEST1 gene function, whether due to a premature stop codon in humans or through MO knockdown in zebrafish, could disrupt critical molecular pathways related to eye, neurological, and skeletal development, leading to similar phenotypes. Further research is required to investigate these mechanisms.
Our study has several limitations. First, the VMD2 manifests with later-onset macular/retinal dystrophy. Thus, we did not examine the retina of zebrafish embryos for macular dystrophy. Second, this is an in vivo knockdown study that validated the essential role of the best1 gene in the early development of zebrafish. Further knockout studies and in vitro experiments are warranted to delineate upstream and downstream targets and pathways involved. Third, the CVJ structures of zebrafish are not as analogical to humans compared with that of mice, which may be more suitable for our future study.
CONCLUSION
Damaging mutation of BEST1 gene may cause a subtype of CVJ malformation with comorbidities such as microphthalmia, VMD2, spinal anomalies, scoliosis, and neurological defects. Further studies are required to investigate the mechanism and possible preventive measures for the subtype of CVJ malformation.









")













